Prolamine Intolerance: Why Gluten Is Only Part Of The Problem
By Jacob Gordon, INHC, FMT-CThis article contains affiliate links. As an Amazon Associate, MyBioHack earns from qualifying purchases at no extra cost to you. We only link products we research and stand behind.
Prolamine intolerance is an immune and digestive reaction to a family of grain storage proteins that extends well beyond wheat gluten.
In this post, we will discuss what prolamines are, which grains contain them, what causes intolerance at a mechanistic level, how prolamines damage the gut and body, which conditions overlap, how to improve tolerance, what to avoid, relevant testing, mechanisms of action, and the genetics involved.

Basics Of Prolamine Intolerance
Prolamines are a class of seed storage proteins found in cereal grains, named for their unusually high content of the amino acids proline and glutamine. R
They are defined by their solubility in alcohol-water mixtures, which distinguishes them from other grain protein fractions like glutelins, albumins, and globulins. R
The most well-known prolamine is gliadin, the toxic fraction of wheat gluten. R
However, "gluten" technically refers to the combination of gliadin (prolamine) and glutenin (glutelin) found only in wheat. R
Every major cereal grain has its own prolamine, and many share structural similarity with wheat gliadin. R
The high proline content makes prolamines resistant to complete breakdown by human digestive enzymes. R
Specifically, the 33-mer peptide from alpha-2-gliadin survives all gastrointestinal proteases, including pepsin, trypsin, chymotrypsin, elastase, and brush border enzymes. R
This 33-amino-acid fragment is the most immunogenic peptide in celiac disease, containing six overlapping T-cell epitopes. R
Celiac disease represents the extreme end of prolamine reactivity, but sensitivity to prolamines exists on a spectrum. R
Non-celiac gluten sensitivity (NCGS) affects an estimated 0.5% to 13% of the population, depending on diagnostic criteria used. R
The problem is broader than gluten itself, because related prolamines in barley, rye, corn, and other grains can trigger similar immune responses. R
Framing the issue as "gluten intolerance" misses the full picture and can lead people to substitute one reactive grain for another. R
Which Grains Contain Prolamines
Each cereal grain has a unique prolamine with a different name, different percentage of total protein, and different degree of immunogenicity.
The following table summarizes the major prolamines across common grains.
| Grain | Prolamine Name | % of Protein | Notes |
|---|---|---|---|
| Wheat | Gliadin | 30-40% | Most studied; contains the 33-mer peptide |
| Barley | Hordein | 35-55% | Cross-reactive with gliadin antibodies |
| Rye | Secalin | 30-50% | Cross-reactive with gliadin antibodies |
| Oats | Avenin | 10-15% | Debated safety; lower prolamine load |
| Corn | Zein | 50-70% | T-cell reactivity in some celiac patients |
| Sorghum | Kafirin | 50-70% | Structurally similar to zein |
| Millet | Panicin/Setarin | 40-60% | Limited immunogenicity data |
| Rice | Oryzin | ~5% | Very low; generally well tolerated |
| Teff | Minimal | <5% | Considered safe for celiac patients |
Corn zein is often overlooked, but research has shown that zein peptides can activate T-cells from celiac patients in vitro, producing interferon-gamma. R
This means corn, which is commonly used as a "gluten-free" replacement, may not be safe for everyone with prolamine sensitivity. R
Oat avenin is debated because while its prolamine content is lower (~10-15%), contamination with wheat during processing is common, and a small subset of celiac patients react to pure avenin itself. R
Pseudocereals like quinoa, amaranth, and buckwheat do not contain prolamines and are generally safe alternatives. R
Rice, with its very low prolamine content (~5%), is typically well tolerated even by celiac patients. R
What Causes Prolamine Intolerance
The root cause is incomplete digestion of proline-rich prolamine sequences, leaving large immunogenic peptide fragments intact in the gut lumen. R
Human gastric and pancreatic proteases lack the ability to cleave at proline residues efficiently, which is why the 33-mer and similar fragments survive. R
Once these intact peptides reach the intestinal epithelium, gliadin binds the CXCR3 receptor on enterocytes, triggering zonulin release via a MyD88-dependent pathway. R
Zonulin is a physiological modulator of tight junctions that, when overproduced, increases intestinal permeability. R
This zonulin-mediated permeability increase occurs in all individuals exposed to gliadin, not only in celiac patients. R
The difference is that celiac and NCGS patients show a more prolonged and exaggerated permeability response compared to healthy controls. R
Amylase-trypsin inhibitors (ATIs), which are non-gluten wheat proteins, independently activate TLR4 on innate immune cells, adding a second inflammatory pathway. R
ATIs represent up to 4% of wheat protein and are potent activators of monocytes, macrophages, and dendritic cells. R
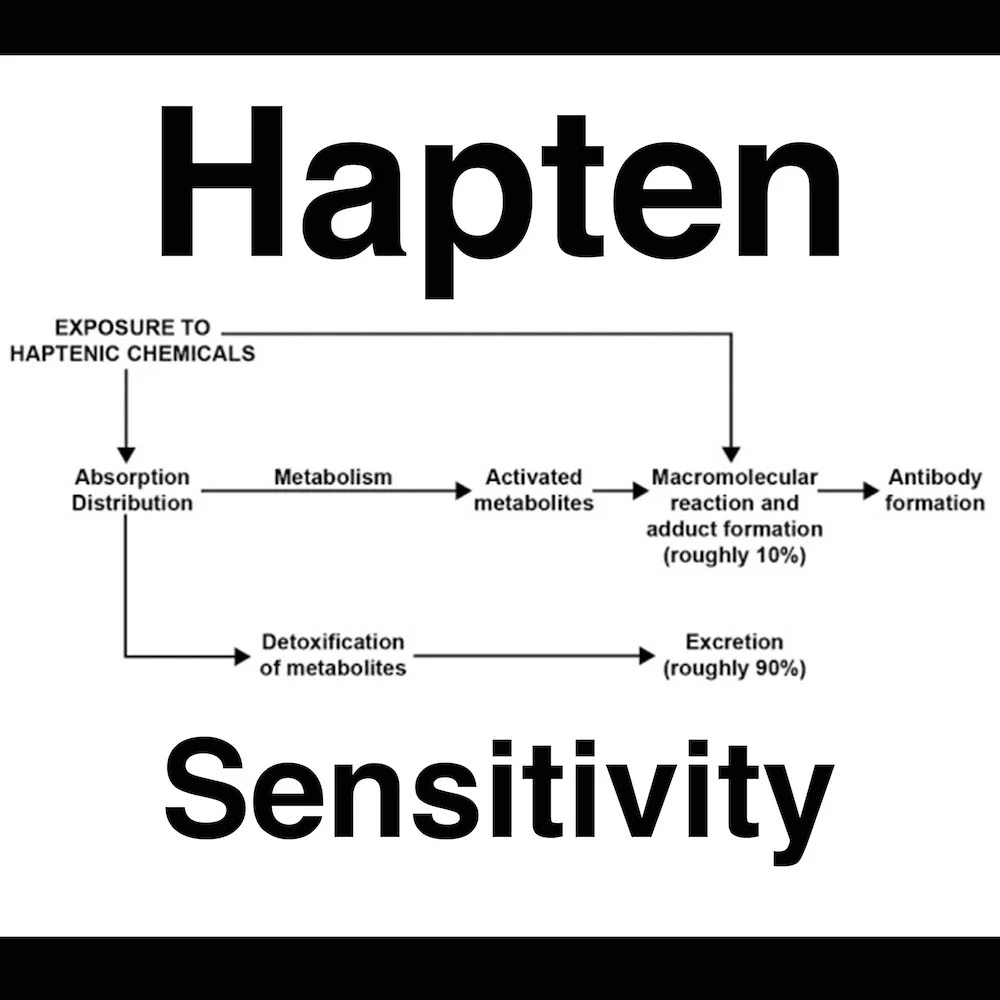
Hapten formation can occur when partially digested prolamine peptides bind to tissue proteins, creating neoepitopes that the immune system recognizes as foreign. R
Modern wheat cultivars may contain higher concentrations of immunogenic epitopes than ancestral varieties, although this is debated. R
Glyphosate exposure has been proposed to worsen prolamine intolerance by disrupting gut microbiome composition and potentially enhancing zonulin release. R
Low secretory IgA (SIgA) production reduces the gut's first-line defense against prolamine peptides at the mucosal surface. R
Gut dysbiosis alters the microbial enzymes that normally assist in partial prolamine degradation, potentially increasing the load of intact immunogenic peptides. R
How Prolamines Damage The Gut And Body
Prolamine-driven damage occurs through multiple overlapping pathways affecting both the gut barrier and systemic immunity.
Tight junction opening via the zonulin pathway increases paracellular permeability, allowing intact prolamine peptides and other luminal antigens (including LPS) to cross into the lamina propria. R
This process is the basis of junction dysfunction, where barrier integrity loss creates a feed-forward cycle of inflammation. R
The glycocalyx, the protective sugar-protein layer lining the intestinal epithelium, can also be degraded by prolamine-driven inflammation, further compromising barrier function. R
Innate immune activation occurs through TLR4 signaling (primarily from ATIs), which activates NF-kB and drives production of IL-1β, IL-6, and TNF-α. R
NCGS appears to be driven primarily by this innate immune pathway, unlike celiac disease which involves both innate and adaptive immunity. R
Mast cell activation in the intestinal mucosa releases histamine, tryptase, and prostaglandins, contributing to local inflammation and visceral hypersensitivity. R
In celiac disease, the enzyme tissue transglutaminase (tTG) deamidates gliadin peptides, converting glutamine to glutamate residues and creating neoepitopes that bind HLA-DQ2/DQ8 with high affinity. R
These deamidated peptides are presented to CD4+ T helper cells, driving an adaptive immune response that produces anti-tTG and anti-deamidated gliadin antibodies. R
IL-15 upregulation in the epithelium activates intraepithelial lymphocytes via the NKG2D receptor, leading to direct killing of enterocytes expressing the stress ligand MICA. R
This IL-15/NKG2D/MICA pathway is responsible for villous atrophy in celiac disease. R
Molecular mimicry between prolamine peptides and human tissue antigens in the brain, thyroid, and pancreas can trigger cross-reactive autoimmune responses. R
The duration of gluten exposure correlates with the risk of developing autoimmune conditions in genetically susceptible individuals. R
Neuroinflammation from cross-reactive antibodies can affect cerebellar Purkinje cells, peripheral nerves, and brain microvasculature. R
Increased intestinal permeability allows bacterial endotoxins (LPS), food antigens, and other inflammatory molecules to enter systemic circulation, creating systemic inflammation. R
Butyrate production by commensal bacteria declines as dysbiosis worsens, further weakening barrier repair. R
NRF2 signaling, which normally activates cytoprotective and antioxidant genes, can be suppressed in the context of chronic prolamine-driven inflammation. R
Prolamine Intolerance And Overlapping Conditions
Prolamine intolerance frequently coexists with or contributes to a wide range of conditions, listed alphabetically.
Autoimmune conditions (Hashimoto's thyroiditis, type 1 diabetes, rheumatoid arthritis) share strong HLA associations with celiac disease, and prolonged gluten exposure increases risk of developing additional autoimmune conditions. R
Dermatitis herpetiformis is the skin manifestation of celiac disease, presenting as intensely itchy, blistering rashes on elbows, knees, and buttocks, driven by IgA deposits in the dermal papillae. R
Eczema and atopic dermatitis have documented associations with wheat sensitivity, with prolamine-driven immune activation contributing to Th2 skewing and skin barrier dysfunction. R
Gluten ataxia and peripheral neuropathy are neurological manifestations caused by anti-gliadin and anti-transglutaminase 6 antibodies targeting cerebellar and peripheral nerve tissue. R
Histamine intolerance and mast cell activation syndrome (MCAS) can be worsened by prolamine-driven intestinal inflammation, which activates mucosal mast cells and increases histamine release. R
Irritable bowel syndrome (IBS) overlaps significantly with NCGS, and gluten has been shown to worsen symptoms in IBS patients even without celiac disease. R
Psoriasis has been linked to anti-gliadin antibodies in a subset of patients, with some studies showing improvement on gluten-free diets. R
SALI (systemic and long-term inflammation) is driven by the persistent intestinal permeability and immune activation that prolamine intolerance creates. R
How To Improve Prolamine Intolerance
Step 1: Strict Elimination (30-90 Days)
Remove all prolamine-containing grains for a minimum of 30 days, ideally 90 days, to allow intestinal healing and immune calming. R
Safe starch alternatives include rice, quinoa, amaranth, buckwheat, sweet potato, white potato, cassava, taro, and plantain. R
Teff is also well tolerated due to its minimal prolamine content. R
Step 2: Enzyme Support For Trace Exposure
DPP-IV Enzyme supplements can help break down trace prolamine peptides from accidental exposure. R
AN-PEP (Aspergillus niger prolyl endoprotease) degrades gluten peptides in the stomach before they reach the small intestine. R
These enzymes are not a substitute for dietary elimination and should only be used as a safety net for inadvertent exposure. R
Step 3: Gut Repair
The following supplements support intestinal barrier restoration and immune regulation (alphabetical).
Colostrum contains immunoglobulins, lactoferrin, and growth factors that support mucosal healing and SIgA production. R
Curcumin inhibits NF-kB activation and reduces TNF-α and IL-6 production in intestinal immune cells. R
L-Glutamine (5-10g per day) is the primary fuel source for enterocytes and supports tight junction protein expression. R
Omega-3 Fish Oil in triglyceride form provides EPA and DHA, which resolve inflammation through specialized pro-resolving mediators (SPMs). R
Probiotics (Lactobacillus rhamnosus GG) modulate intestinal immune responses and enhance barrier function. R
Quercetin stabilizes mast cells and reduces histamine release from prolamine-driven mast cell activation. R
Vitamin D3 + K2 supports immune regulation, tight junction integrity, and mucosal vitamin D signaling. R
Zinc Carnosine has been shown to increase villous height, reduce gastric permeability, and stabilize the gut lining. R
Step 4: Reintroduction Hierarchy
After the elimination period, reintroduce grains one at a time, starting with the lowest-risk options and progressing toward higher-prolamine grains.
A suggested order (from safest to most reactive): rice, certified gluten-free oats, corn, sorghum, millet, rye, barley, and wheat last. R
Wait at least 3 days between introducing each grain to monitor for delayed reactions. R
Track symptoms including digestive changes, skin reactions, brain fog, joint pain, and fatigue. R
What To Stay Away From
During the elimination phase, avoid all of the following.
All wheat products (bread, pasta, couscous, flour tortillas, crackers, pastries, and baked goods). R
Barley, rye, spelt, kamut, triticale, farro, and einkorn. R
Hidden gluten sources including soy sauce, malt extract, malt vinegar, beer, seitan, imitation crab, and many processed foods. R
Medications and supplements that use wheat starch or gluten-containing binders as excipients. R
NSAIDs (ibuprofen, aspirin, naproxen) increase intestinal permeability independently and amplify prolamine-driven barrier disruption. R
Excessive alcohol disrupts tight junctions and increases intestinal permeability through acetaldehyde-mediated damage. R
Chronic psychological stress elevates cortisol, which increases intestinal permeability through CRH-mediated mast cell activation. R
Intense, prolonged exercise causes transient barrier disruption through splanchnic hypoperfusion and can worsen prolamine sensitivity during active inflammation. R
Emulsifiers in processed foods (polysorbate 80, carboxymethylcellulose) can disrupt the mucosal barrier and alter gut microbiome composition. R
Biofilm-forming organisms in the gut may harbor prolamine peptides and perpetuate immune activation even during dietary elimination. R
Testing
Blood Markers
tTG IgA (tissue transglutaminase IgA) is the primary screening test for celiac disease, with sensitivity above 95% in active disease. R
I use the tTG IgA (Quest) test for initial celiac screening.
Deamidated gliadin peptide (DGP) IgA and IgG captures celiac patients who are IgA-deficient (2-3% of celiac patients have selective IgA deficiency). R
I use the DGP IgA (Quest) test for supplementary celiac screening.
Total IgA should be measured alongside tTG IgA, because IgA deficiency will produce a false negative. R
Anti-gliadin IgA and IgG are older markers that are less specific for celiac but can indicate NCGS when celiac has been ruled out. R
Comprehensive Panels
I use the Wheat Antibodies (Vibrant Wellness) panel for a detailed wheat protein reactivity profile covering gliadin, glutenin, gluteomorphin, wheat germ agglutinin, and deamidated gliadin. R
I use the Lectin Antibodies (Vibrant Wellness) panel to assess reactivity to lectins across grains and legumes.
I use the Corn Antibodies (Vibrant Wellness) panel to check for zein and other corn protein reactivity, which is missed by standard celiac screening. R
I use the Food Sensitivity Complete (Vibrant Wellness) for a broad food reactivity screen covering IgG and IgA to over 200 foods.
I use the Food Antibodies Bundle (All 7) (Vibrant Wellness) for the most comprehensive food reactivity assessment, covering wheat, corn, dairy, egg, lectin, seafood, and nut panels.
I use the Gut Zoomer (Vibrant Wellness) to assess intestinal permeability markers (zonulin, LPS antibodies, actin antibodies), microbiome composition, and pathogen screening.
Celiac Diagnostic Panel
I use the Celiac Disease Diagnostic Panel (Quest) as a comprehensive celiac workup including tTG IgA, DGP IgA/IgG, and total IgA in a single panel.
Genetics
I use the Celiac Genetics (Vibrant Wellness) panel to check HLA-DQ2 and HLA-DQ8 status. R
I use the HLA Typing for Celiac Disease (Quest) as an alternative HLA typing option.
HLA-DQ2/DQ8 is necessary but not sufficient for celiac disease, so a negative result effectively rules out celiac, while a positive result indicates susceptibility. R
Immune And Inflammation Panels
I use the Immune Zoomer (Vibrant Wellness) to assess autoantibodies and immune markers that may indicate prolamine-triggered autoimmunity.
I use the Foundation Zoomer (Vibrant Wellness) as a baseline metabolic and inflammatory panel.
Elimination Diet
The elimination diet remains the gold standard for diagnosing non-celiac prolamine sensitivity, because no single blood test reliably identifies it. R
The Salerno criteria recommend a double-blind, placebo-controlled crossover challenge for NCGS diagnosis in research settings. R
In clinical practice, a structured elimination followed by systematic reintroduction with symptom tracking is the most practical approach. R
Mechanisms Of Action
Simple
Prolamines resist human digestive enzymes because of their high proline content, leaving large peptide fragments intact in the gut.
These intact peptides force open the tight junctions between gut lining cells, allowing them and other inflammatory molecules to leak through.
Once through the gut wall, prolamine peptides activate the immune system, triggering inflammation in the gut, brain, skin, and other organs.
In genetically susceptible individuals, the immune system begins attacking the body's own tissues through antibody cross-reactivity.
Advanced
Zonulin/CXCR3/EGFR/PAR2 Pathway
Gliadin peptides bind CXCR3 on the apical surface of enterocytes, initiating a MyD88-dependent signaling cascade. R
This triggers zonulin (pre-haptoglobin 2) release, which acts on EGFR and PAR2 receptors on the basolateral surface. R
EGFR/PAR2 activation phosphorylates zonula occludens proteins (ZO-1), displacing them from tight junction complexes. R
This leads to disassembly of claudin and occludin from the tight junction, increasing paracellular permeability. R
The response is dose-dependent, reversible in healthy individuals, but sustained and exaggerated in celiac and NCGS patients. R
TLR4/MyD88/NF-kB Pathway (ATIs)
Amylase-trypsin inhibitors activate TLR4 on monocytes, macrophages, and dendritic cells in the intestinal lamina propria. R
TLR4 engagement recruits the adaptor protein MyD88, activating IRAK4, TRAF6, and the IKK complex. R
The IKK complex phosphorylates IkBα, releasing NF-kB to translocate to the nucleus and drive transcription of IL-1β, IL-6, IL-8, and TNF-α. R
This pathway operates independently of the adaptive immune response and is the primary driver of innate inflammation in NCGS. R
tTG Deamidation And Neoepitope Formation
Tissue transglutaminase (tTG2) catalyzes the deamidation of glutamine residues to glutamic acid within gliadin peptides. R
This converts neutral glutamine to negatively charged glutamate, dramatically increasing binding affinity for the HLA-DQ2 and HLA-DQ8 peptide-binding groove. R
Antigen-presenting cells display deamidated peptides to CD4+ T-cells, which produce IFN-γ and drive B-cell production of anti-tTG and anti-DGP antibodies. R
Anti-tTG antibodies have direct pathogenic effects on the epithelium, independent of T-cell activity. R
IL-15/NKG2D/MICA Pathway
Gliadin-driven epithelial stress increases IL-15 expression in the intestinal epithelium. R
IL-15 upregulates the activating receptor NKG2D on intraepithelial CD8+ T-cells and natural killer cells. R
Simultaneously, stressed enterocytes express the NKG2D ligand MICA on their surface. R
NKG2D-MICA interaction triggers direct cytolysis of enterocytes by intraepithelial lymphocytes, independent of antigen-specific T-cell receptor signaling. R
This pathway is responsible for villous atrophy and crypt hyperplasia, the hallmark histological findings of celiac disease. R
Genetics
HLA-DQ2.5
The haplotype DQA1*05:01 / DQB1*02:01 encodes the HLA-DQ2.5 heterodimer. R
Approximately 90-95% of celiac disease patients carry at least one copy of HLA-DQ2.5. R
The DQ2.5 peptide-binding groove has a strong preference for negatively charged (deamidated) prolamine peptides at anchor positions P4, P6, and P7. R
Homozygosity for DQB1*02 (encoding DQ2.5 on both chromosomes) carries the highest celiac risk, approximately fivefold higher than heterozygosity. R
HLA-DQ8
The haplotype DQA1*03:01 / DQB1*03:02 encodes the HLA-DQ8 heterodimer. R
Approximately 5-10% of celiac disease patients carry HLA-DQ8 without DQ2.5. R
DQ8 preferentially binds peptides with a positive charge at P1 and hydrophobic residues at P9. R
DQ8 is also associated with type 1 diabetes susceptibility, creating a shared genetic link between the two autoimmune conditions. R
IL2/IL21 (4q27)
The IL2/IL21 locus on chromosome 4q27 is the strongest non-HLA genetic association with celiac disease. R
IL-21 drives proliferation of gliadin-reactive T-cells and promotes IFN-γ production in the intestinal mucosa. R
This locus is shared with other autoimmune diseases including type 1 diabetes and rheumatoid arthritis. R
MYO9B
MYO9B encodes myosin IXB, an unconventional myosin involved in tight junction regulation and epithelial barrier integrity. R
Variants in MYO9B have been associated with celiac disease susceptibility and increased intestinal permeability. R
MYO9B regulates RhoA GTPase signaling, which controls actin cytoskeleton dynamics at tight junctions. R
TLR4 Variants
TLR4 polymorphisms can alter the innate immune response to ATIs and other microbial ligands in the gut. R
The Asp299Gly (rs4986790) and Thr399Ile (rs4986791) variants reduce TLR4 responsiveness and have been associated with altered inflammatory bowel disease susceptibility. R
Individuals with gain-of-function TLR4 variants may have exaggerated innate responses to ATIs. R
Other Relevant Loci
A GWAS meta-analysis identified 39 non-HLA loci associated with celiac disease, with the majority involved in immune regulation. R
These include genes related to T-cell differentiation (TAGAP, SH2B3), innate immunity (REL, UBE2L3), and B-cell signaling (TNFRSF14). R
Many celiac risk loci overlap with susceptibility loci for other autoimmune conditions, explaining the high rate of autoimmune comorbidity. R
Zinc and copper metabolism genes may also influence prolamine tolerance through their effects on intestinal barrier function and immune regulation. R
More Research
Gliadin exposure increases intestinal permeability in all individuals tested, not only those with celiac disease, through zonulin-dependent mechanisms. R
Corn zein prolamines induce T-cell reactivity and interferon-gamma production in celiac patients, suggesting that corn may not be a universally safe substitute in gluten-free diets. R
Non-celiac wheat sensitivity involves innate immune activation without the adaptive immune hallmarks (anti-tTG antibodies, villous atrophy) seen in celiac disease. R
Sourdough fermentation using specific Lactobacillus strains can partially degrade gluten peptides, including the 33-mer, reducing immunogenicity (although residual reactivity may persist). R
A comprehensive review of grains and inflammation found that prolamine-driven permeability increases, lectin binding, and inhibition of nutrient absorption are shared mechanisms across multiple grain types. R
Gluten triggers gastrointestinal symptoms in a significant subset of IBS patients, with symptoms reproducible on blinded rechallenge. R
Gluten ataxia represents a form of sporadic cerebellar ataxia caused by anti-gliadin antibodies cross-reacting with cerebellar Purkinje cell epitopes, with improvement documented on strict gluten-free diet. R
The duration of gluten exposure before diagnosis of celiac disease correlates with the risk of developing additional autoimmune disorders, emphasizing the importance of early identification and elimination. R
Jacob Gordon
INHC, FMT-C
Board Certified Health Coach
I spent years battling unexplained chronic illness before discovering biohacking, epigenetics, and functional medicine. Now I share that research at MyBioHack to help others find their own answers.
Book a ConsultationRelated Protocols & Supplements
Deep-dive chapters and recommended supplements for this topic
Quercetin
500mg 2x/day
Vitamin D3 + K2
5000 IU + 200mcg/day
DAO Enzyme
1 cap before meals